
![]()
![]()
![]()
February 20, 1970
![]()
of the Leptodactylid Frog Genus
Syrrhophus Cope
Lawrence
1970
Editors of this number:
Frank B. Cross, Philip S. Humphrey, William E. Duellman
Volume 20, No. 1, pp. 1-45, 22 figs.
Published February 20, 1970
University of Kansas
Lawrence, Kansas
THE UNIVERSITY OF KANSAS PRINTING SERVICE
LAWRENCE, KANSAS
1970
Genus Syrrhophus Cope
Cope (1878) proposed the genus Syrrhophus for a medium-sized
leptodactylid frog from central Texas; in the ensuing 75 years the genus
was expanded to include a heterogeneous group of frogs ranging from
Texas to Peru. Taylor (1952) and Firschein (1954) limited the genus to
several species of frogs occurring in Guatemala, México, and Texas.
Lynch (1968) provided a definition of the previously loosely-defined
genus.
With the exception of Taylor (1952), who treated the Costa Rican
species, none of these authors dealt with the present status of the
nineteen species erroneously assigned to Syrrhophus. These species are
listed in Tables 1 and 2 with the name currently applied. Some of them
are new combinations and their justifications will be published
elsewhere. Gorham (1966) is the most recent author to include South
American species in the genus Syrrhophus.
Smith and Taylor (1948) recognized two species groups of the genus in
México, an eastern and a western group (here termed complexes for
purposes of discussion), separated on the basis of the number of palmar
(metacarpal) tubercles (three palmar tubercles in the members of the
eastern complex and two in those of the western complex). Duellman
(1958) reviewed the species of the genus occurring in western México and
concluded that there were five species (two polytypic). Dixon and Webb
(1966) described an additional species from Jalisco, México. The
distributions of some species have been extended, but
otherwise the western complex of species remains unchanged since
Duellman’s review.
Smith and Taylor (1948) recognized seven species of the genus in eastern
México. Firschein revised the eastern complex (as then understood), and
in so doing added one new species and treated Syrrhophus verruculatus
as a nomen dubium. Dixon (1957) redefined the related genus
Tomodactylus and transferred T. macrotympanum Taylor to the genus
Syrrhophus. Neill (1965) described a new subspecies of S. leprus
from British Honduras. Two species (S. gaigeae and S. marnockii)
were recognized in Texas until Milstead, Mecham, and McClintock (1950)
synonymized S. gaigeae
[Pg 4]
with S. marnockii. Thus, at present, nine
species (one polytypic) are recognized on the eastern slopes and
lowlands from central Texas to British Honduras. These are currently
placed on one species group equivalent to the western complex reviewed
by Duellman (1958).
Table 1—Species Described as Members of the Genus Syrrhophus
but Now Placed in Other Genera.
| Trivial name and author | Current combination | |
| areolatus Boulenger, 1898 | Eleutherodactylus areolatus | |
| calcaratus Andersson, 1945 | Eleutherodactylus anderssoni | |
| caryophyllaceus Barbour, 1928 | Eleutherodactylus caryophyllaceus | |
| coeruleus Andersson, 1945 | Eleutherodactylus coeruleus | |
| ineptus Barbour, 1928 | Eleutherodactylus diastema | |
| juninensis Shreve, 1938 | Eupsophus juninensis | |
| lutosus Barbour and Dunn, 1921 | Eleutherodactylus lutosus | |
| molinoi Barbour, 1928 | Eleutherodactylus molinoi | |
| montium Shreve, 1938 | Niceforonia montia | |
| mystaceus Barbour, 1922 | Eleutherodactylus rhodopis | |
| obesus Barbour, 1928 | Eleutherodactylus punctariolus | |
| omiltemanus Gunther, 1900 | Eleutherodactylus omiltemanus[1] | |
| pardalis Barbour, 1928 | Eleutherodactylus pardalis |
Table 2—Species Incorrectly Regarded as Members of the Genus Syrrhophus
but Described as Members of Other Genera.
| Trivial name, original generic assignment, and author | Current combination | |
| chalceus (Phyllobates) Peters, 1873 | Eleutherodactylus chalceus | |
| festae (Paludicola) Peracca, 1904 | Niceforonia festae | |
| hylaeformis (Phyllobates) Cope, 1875 | Eleutherodactylus hylaeformis | |
| palmatus (Phyllobates) Werner, 1899 | Colostethus palmatus | |
| ridens (Phyllobates) Cope, 1866 | Eleutherodactylus ridens | |
| simonsii (Paludicola) Boulenger, 1900 | Niceforonia simonsii |
Table 3—Nominal Species of Syrrhophus (sensu strictu)
and the Name Used Herein.
| Original combination | Current combination | |
| campi, Syrrhophus | cystignathoides campi | |
| cholorum, Syrrhophus leprus | leprus | |
| cystigathoides, Phyllobates | cystignathoides cystignathoides | |
| dennisi, Syrrhophus | dennisi new species | |
| gaigeae, Syrrhophus | guttilatus | |
| guttilatus, Malachylodes | guttilatus | |
| interorbitalis, Syrrhophus | interorbitalis | |
| latodactylus, Syrrhophus | longipes | |
| leprus, Syrrhophus | leprus | |
| longipes, Batrachyla | longipes | |
| macrotympanum, Tomodactylus | verrucipes | |
| marnockii, Syrrhophus | marnockii | |
| modestus, Syrrhophus | modestus | |
| nebulosus, Syrrhophus | pipilans nebulosus | |
| nivocolimae, Syrrhophus | nivocolimae | |
| pallidus, Syrrhophus modestus | pallidus | |
| petrophilus, Syrrhophus | guttilatus | |
| pipilans, Syrrhophus | pipilans pipilans | |
| rubrimaculatus, Syrrhophus | rubrimaculatus | |
| smithi, Syrrhophus | guttilatus | |
| teretistes, Syrrhophus | teretistes | |
| verrucipes, Syrrhophus | verrucipes | |
| verruculatus, Phyllobates | Nomen dubium |
[Pg 5]
In the course of preparing an account of the species of
Eleutherodactylus occurring in México and northern Central America, it
became necessary to reëxamine the status of the genus Syrrhophus and
its nominal species. It soon became evident that there were more names
than species, that some previously regarded species were geographic
variants, and that the eastern and western groups (complexes here) were
artificial divisions of the genus. I conclude that there are seven
species (one polytypic)
of Syrrhophus in eastern México,
Texas, and El Petén of Guatemala, and seven species (one polytypic) in
western México. The current status of each of the 23 names correctly
assigned to the genus is presented in Table 3.
The fourteen species recognized by me are placed in five species groups.
Two of these groups are presently placed in the western complex
(modestus and pipilans groups) and three in the eastern complex
(leprus, longipes and marnockii groups). The two complexes do not
correspond exactly with the eastern and western groups of Smith and
Taylor (1948), Firschein (1954), and Duellman (1958) since S.
rubrimaculatus is now associated with the eastern leprus group.
The definitions and contents of the five species groups are as follows:
in outline; first finger longer or shorter than second; snout
acuminate or subacuminate, not rounded; outer metatarsal tubercle
conical; digits lacking distinct lateral fringes.
content: cystignathoides, leprus and rubrimaculatus.
first finger shorter than second; snout acuminate; outer metatarsal
tubercle not conical; digits bearing lateral fringes.
content: dennisi and longipes.
outline; first finger equal in length to second or slightly shorter;
snout rounded; outer metatarsal tubercle not conical; digits lacking
lateral fringes; generally stout-bodied frogs.
content: guttilatus, marnockii, and verrucipes.
second fingers subequal in length, first usually slightly shorter than
second; snout subacuminate; inner metatarsal tubercle twice as large (or
larger) as outer metatarsal tubercle; digits bearing poorly-defined
lateral fringes.
content: interorbitalis, modestus, nivocolimae, pallidus,
and teretistes.
truncate in outline; first finger equal in length to second; snout
subacuminate; metatarsal tubercles subequal in size; digits lacking
lateral fringes.
content: pipilans.
Acknowledgments.—For loan of specimens, I am indebted to Richard
J. Baldauf, Texas A & M University (TCWC); W. Frank Blair,
University of Texas (TNHC); Charles M. Bogert and Richard G.
Zweifel, American Museum of Natural History (AMNH); James E. Böhlke
and Edmond V. Malnate, Academy of Natural Sciences of Philadelphia
(ANSP); Robert F. Inger and Hymen Marx, Field Museum of Natural
History (FMNH); Ernest A. Liner (EAL); Michael Ovchynnyk, Michigan
State University collection (MSU); James A. Peters, United States
National Museum (USNM); Douglas A. Rossman, Louisiana State
University Museum of Zoology (LSUMZ); Hobart M. Smith, University
of Illinois Museum of Natural History (UIMNH); Charles F. Walker,
University of Michigan Museum of Zoology (UMMZ); and John W.
Wright, Los Angeles County Museum (LACM). Specimens in the
collection at the University of Kansas Museum of Natural History
are identified as KU. The abbreviations EHT-HMS refer to the Edward
H. Taylor-Hobart M. Smith collection and FAS to the Frederick A.
Shannon collection. The type-specimens from these collections are
now in the Field Museum of Natural History and the University of
Illinois Museum of Natural History.
I have profited from discussions concerning this problem with
several persons, most notably William E. Duellman, Hobart M. Smith,
Edward H. Taylor and Charles F. Walker. Nevertheless, the ideas and
conclusions presented here should not be construed as necessarily
reflecting their opinions.
David M. Dennis executed all of the figures, and my wife, Marsha,
typed the manuscript.
Materials and Methods.—In the course of this study, 1003 specimens
of the genus were examined. The holotypes of 21 of the 23
nominal species are extant; I have examined 19 of these. Nine
measurements were taken, and five ratios computed for each of 338
specimens. Females are available for all species but one; thus,
measurements were taken on individuals of both sexes.
Size and proportions.—Frogs of this genus range in size from 16 to 40
mm. in snout-vent length. Five species are relatively small: S.
cystignathoides, modestus, nivocolimae, pallidus and
rubrimaculatus; one, S. longipes, is relatively large, and the
remaining eight species are intermediate in size (22-30 mm.).
Males are generally smaller than females and have proportionately longer
heads and usually larger tympani. No significant differences were found
among proportions, except that S. longipes has a larger tympanum/eye
ratio than any other species. Frogs in the Syrrhophus marnockii group
tend to have shorter shanks and feet, thereby giving those species a
more stocky appearance. However, the differences are not significant.
A summary of the data on size and proportions for the frogs of the genus
Syrrhophus is given in Tables 4, 5, and 6.
[Pg 7]
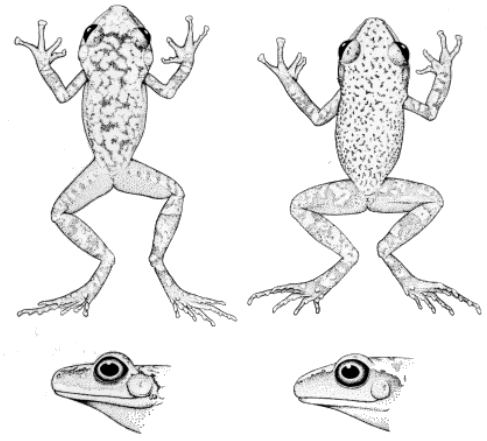
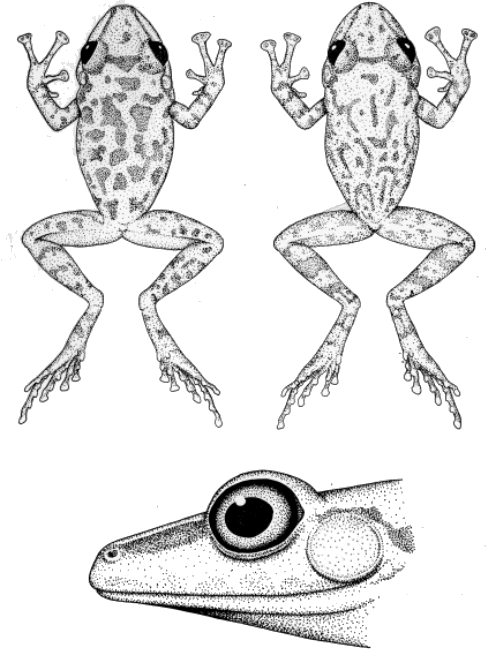
Hands and Feet.—Taylor and Smith (1945), Smith and Taylor (1948),
Firschein (1954) and Duellman (1958) discussed the value of the palmar
tubercles in identifying frogs of this genus. The eastern complex in
general has a well-developed outer palmar tubercle (Fig. 1) in
distinction to the western complex in which the outer palmar tubercle is
reduced or absent (Fig. 2). Dixon and Webb (1966) imply that the outer
palmar tubercle is rarely absent but is usually smaller than the first
supernumerary tubercle of the fourth finger. My study of the western
species demonstrates that the outer palmar tubercle is indeed usually
present and smaller than the first supernumerary tubercle.
Differences in interpretation of the terms “unexpanded” and “narrow,” as
well as differences in techniques of preservation, have led to confusion
of the reported digital shapes in various species. Constant specific
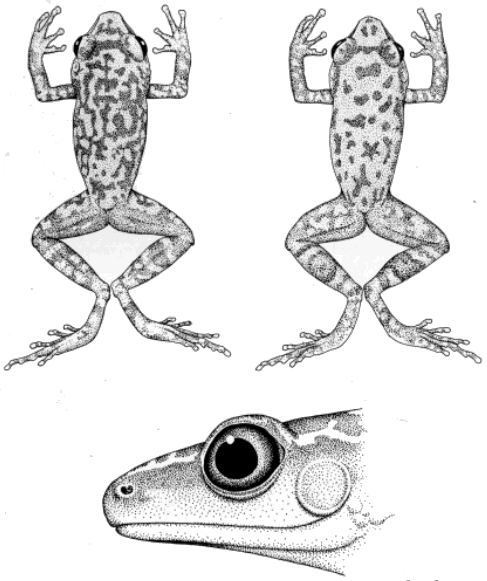
differences are evident in the hands (Fig. 1). Except in the cases of
excessive uptake of fluids, all species have a terminal transverse
groove at the tip of each digit. Taylor (1940b) stated that S. smithi
lacked grooves, but examination of the holotype reveals faint grooves at
the tops of the digits. Syrrhophus guttilatus, leprus, pipilans,
and verrucipes lack lateral fringes on the fingers. Lateral fringes
are well developed in the longipes and modestus groups but poorly
defined or absent in the other members of the genus. The digital pads of
the frogs of the longipes group are much broader than those of the
other species and are narrowest in the frogs of the leprus group.
Supernumerary tubercles are present on the palmar surfaces of all
species of the genus.
Table 4—Size and Proportions in the Frogs
of the Syrrhophus leprus Group.
| Species | Sex | N | Snout-vent length (SVL) | Tibia length/ SVL | Head width/ SVL | Tympanum/ Eye | Eyelid/ Interorbital |
| cystignathoides campi | ♂ | 33 | 16.3-23.5 | 41.3-49.6 | 34.0-40.1 | 43.7-66.5 | 43.2-89.6 |
| (45.8) | (37.0) | (56.2) | (61.5) | ||||
| ♀ | 12 | 16.0-25.8 | 41.5-51.0 | 33.0-38.0 | 42.8-60.0 | 48.2-69.2 | |
| (45.8) | (35.0) | (51.2) | (60.1) | ||||
| c. cystignathoides | ♂ | 15 | 16.8-22.1 | 45.1-50.4 | 33.2-40.7 | 44.3-68.7 | 44.6-65.4 |
| (47.3) | (37.8) | (54.8) | (60.0) | ||||
| ♀ | 6 | 19.6-24.2 | 46.4-50.0 | 34.1-38.1 | 43.3-56.5 | 53.2-65.4 | |
| (47.6) | (36.2) | (46.9) | (59.2) | ||||
| leprus | ♂ | 14 | 20.6-26.4 | 42.3-52.3 | 35.0-40.3 | 47.5-62.5 | 58.2-72.5 |
| (46.8) | (37.4) | (56.5) | (67.3) | ||||
| ♀ | 15 | 22.1-29.2 | 43.4-53.3 | 32.6-38.9 | 38.6-57.9 | 50.2-86.9 | |
| (47.1) | (35.8) | (47.1) | (68.1) | ||||
| rubrimaculatus | ♂ | 12 | 18.2-23.5 | 40.4-46.2 | 31.8-35.5 | 35.5-46.5 | 65.1-78.5 |
| (43.4) | (33.8) | (41.7) | (71.7) |
Table 5—Size and Proportions in the Frogs
of the Syrrhophus longipes and S. marnockii Groups.
| Species | Sex | N | Snout-vent length (SVL) | Tibia length/ SVL | Head width/ SVL | Tympanum/ Eye | Eyelid/ Interorbital |
| dennisi | ♂ | 16 | 22.8-28.4 | 43.9-49.7 | 35.3-41.2 | 53.9-64.2 | 55.3-74.0 |
| (47.4) | (38.8) | (58.9) | (65.1) | ||||
| ♀ | 10 | 25.9-32.0 | 46.3-50.8 | 35.6-40.3 | 50.6-58.7 | 58.1-70.9 | |
| (48.2) | (37.7) | (54.9) | (63.6) | ||||
| longipes | ♂ | 22 | 22.1-33.2 | 45.8-51.7 | 38.7-44.4 | 61.1-87.2 | 61.5-83.0 |
| (48.4) | (41.8) | (72.0) | (72.0) | ||||
| ♀ | 19 | 26.8-39.6 | 44.3-51.0 | 36.3-40.8 | 49.5-72.1 | 55.3-85.9 | |
| (47.2) | (39.1) | (59.5) | (67.9) | ||||
| guttilatus | ♂ | 19 | 20.6-29.0 | 41.2-48.1 | 36.9-44.9 | 55.1-75.7 | 53.3-79.5 |
| (44.5) | (40.6) | (64.1) | (66.0) | ||||
| ♀ | 5 | 25.7-31.0 | 41.4-46.8 | 35.9-42.3 | 47.6-61.7 | 62.3-79.8 | |
| (43.6) | (38.5) | (54.0) | (72.9) | ||||
| marnockii | ♂ | 14 | 18.4-28.9 | 42.3-47.2 | 36.1-43.0 | 47.2-68.3 | 51.6-74.4 |
| (44.1) | (39.6) | (61.2) | (66.3) | ||||
| ♀ | 29 | 20.4-35.4 | 38.7-46.4 | 35.9-41.3 | 45.8-73.3 | 52.1-70.5 | |
| (42.7) | (38.2) | (60.3) | (60.7) | ||||
| verrucipes | ♂ | 29 | 17.5-29.2 | 42.7-49.5 | 36.2-42.4 | 56.1-82.2 | 56.8-82.8 |
| (46.3) | (39.1) | (67.8) | (70.4) | ||||
| ♀ | 6 | 26.5-31.7 | 42.4-47.7 | 36.0-38.1 | 45.8-57.8 | 61.0-77.9 | |
| (44.6) | (37.0) | (53.9) | (69.0) |
Table 6—Size and Proportions in the Frogs
of the Syrrhophus pipilans and S. modestus Groups.
| Species | Sex | N | Snout-vent length (SVL) | Tibia length/ SVL | Head width/ SVL | Tympanum/ Eye | Eyelid/ Interorbital |
| pipilans nebulosus | ♂ | 17 | 22.9-28.5 | 38.1-42.0 | 34.4-37.2 | 36.6-47.8 | 56.1-82.4 |
| ♀ | ♀ | ♀ | (40.0) | (35.4) | (43.6) | (68.2) | |
| ♀ | 3 | 21.1-22.7 | 42.1-44.5 | 33.2-35.8 | 36.6-47.6 | 64.3-65.4 | |
| pipilans pipilans | ♂ | 18 | 22.6-27.8 | 37.9-44.0 | 32.2-36.5 | 38.0-54.0 | 56.1-79.5 |
| (41.4) | (33.0) | (46.2) | (67.3) | ||||
| ♀ | 1 | 29.4 | 38.4 | 32.5 | 44.6 | 55.0 | |
| modestus | ♂ | 8 | 15.8-20.1 | 38.5-42.6 | 32.1-38.1 | 26.8-39.3 | 57.0-86.9 |
| (40.6) | (34.2) | (31.5) | (69.1) | ||||
| ♀ | 1 | 18.5 | 44.2 | 36.0 | 24.0 | 52.1 | |
| pallidus | ♂ | 6 | 17.9-19.3 | 41.0-44.9 | 32.6-36.2 | 27.0-35.6 | 59.4-67.7 |
| (43.4) | (35.2) | (30.9) | (65.2) | ||||
| teretistes | ♂ | 18 | 19.2-23.2 | 41.5-45.3 | 32.5-36.4 | 28.6-43.8 | 51.2-75.0 |
| (43.7) | (34.0) | (33.7) | (62.2) | ||||
| ♀ | 1 | 24.8 | 41.8 | 30.8 | 37.9 | 60.5 | |
| nivocolimae | ♂ | 15 | 18.9-21.1 | 42.2-48.6 | 30.9-37.1 | 30.0-39.3 | 42.6-69.1 |
| (45.0) | (33.7) | (34.7) | (55.0) | ||||
| ♀ | 1 | 24.1 | 40.9 | 33.5 | 27.6 | 56.5 | |
| interorbitalis | ♂ | 1 | 25.6 | 43.0 | —— | 39.4 | 57.6 |
| ♀ | 9 | 20.2-26.7 | 39.9-47.1 | 32.6-39.3 | 29.1-41.2 | 58.2-76.9 | |
| (43.2) | (35.8) | (36.4) | (69.2) |
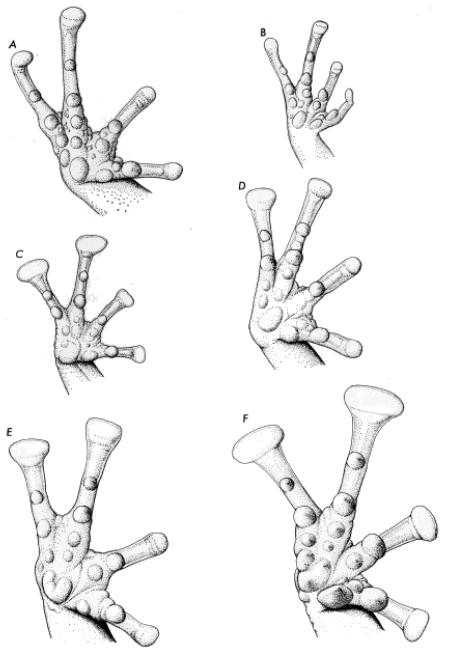
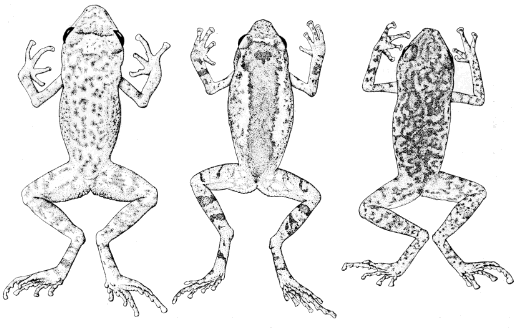
Syrrhophus. (A) verrucipes (UIMNH 15995), (B) rubrimaculatus (KU
58911), (C) dennisi sp. nov. (holotype, UMMZ 101121), (D) guttilatus
(UIMNH 55520), (E) marnockii (TCWC 4782), and (F) longipes (TCWC
12179). All ×6.5.
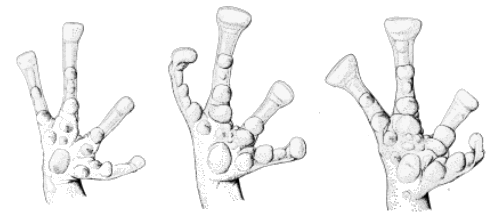
Syrrhophus. pipilans (left, KU 58908, ×6) and teretistes (center,
KU 75269, and right, KU 75263, respectively, ×9).
In S. cystignathoides and leprus, the first finger is longer than
the second, and the first two fingers are equal in length in
guttilatus and marnockii. In the other species the first finger is
shorter than the second.
Supernumerary tubercles are well developed on the plantar surfaces in
all species, except S. guttilatus, in which they are poorly defined
(Fig. 3). The relative sizes of the metatarsal tubercles has been used
in the classification of the species and species groups of Syrrhophus.
The metatarsal tubercles are similar in all species of the eastern
complex (including rubrimaculatus); the outer tubercle is always about
one-half the size of the ovoid inner metatarsal tubercle. In the
leprus group the outer tubercle is conical and compressed. The
metatarsal tubercles of pipilans are about the same size, or the outer
is slightly smaller than the inner. In the modestus group the outer
metatarsal tubercle is about one-third the size of the inner.
All species, except guttilatus, have well-defined to poorly defined
lateral fringes on the toes. All species have expanded toe pads. The
fifth toe is usually shorter than the third, but the second is equal in
length to the fifth in some specimens of S. cystignathoides and S.
marnockii. Syrrhophus nivocolimae is the only species with tubercles
along the outer edge of the tarsus; this is merely a reflection of the
highly tuberculate nature of the skin in this species.
Skin texture.—The skin of the dorsum is smooth or very weakly
pustular in all species of the genus except nivocolimae and
verrucipes. The dorsal surfaces of nivocolimae are warty; in
verrucipes the skin is pustular. The skin of the venter is areolate in
cystignathoides
[Pg 11]
cystignathoides, dennisi and verrucipes but is
smooth in all other species of the genus.
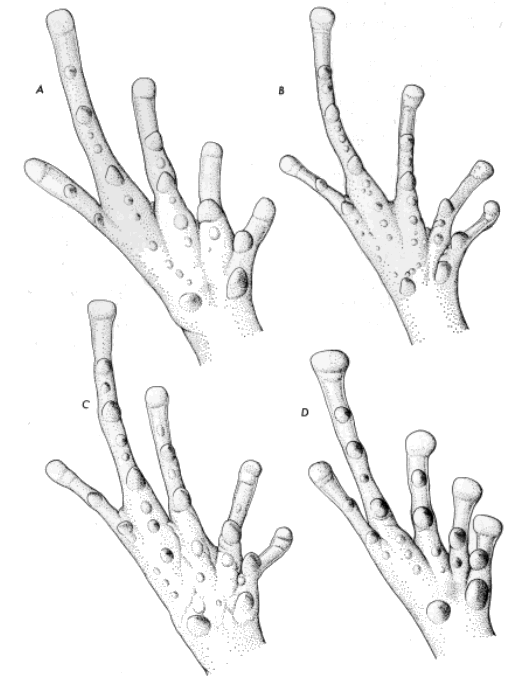
Syrrhophus. (A) guttilatus (UIMNH 55519, ×6), (B) leprus (UIMNH
42726, ×6), (C) verrucipes (UIMNH 15995, ×6), and (D) longipes (TCWC
12179, ×4.6).
Color pattern.—As is evident in the diagnoses, the color patterns
of given populations have been regarded as useful in separating the
[Pg 12]
species and subspecies. Duellman (1958) suggested that the coloration,
with the exception of modestus, was a dark ground color with
pale markings. It is a moot point whether the frogs have light spots
on a dark background or have a light background with an extensive
reticulate dark pattern. The venters are gray or white, and the
vocal sac is nearly black in some species. Interorbital dark bars or
triangles are absent in only two species of the eastern complex,
cystignathoides campi and marnockii; the latter lacks a supratympanic
stripe, which is present in the other members of the eastern
complex. Syrrhophus interorbitalis and nivocolimae have light
interorbital
bars; these bars occur in only one other population of the
genus (S. c. cystignathoides). Bars on the thighs are ill defined or
absent in the members of the marnockii and part of the modestus
groups. The color in life is noted in the species accounts.
Voice.—The voices of all Syrrhophus can be described as a
single short chirp or peep; without audiospectrographic analyses
the significance of the differences between a chirp, peep, or short
whistle cannot be appreciated. Martin (1958) and Wright and
Wright (1949) reported multi-noted calls, and one collector of S.
verrucipes noted the frog “trilled.”
Fouquette (1960) presented analyses of two species (marnockii
and pipilans nebulosus). The voices were very similar; both frogs
were reported to “trill” and “chirp.”
The genus Syrrhophus has been defined (Lynch, 1968) and
limited to the group of species occurring in Guatemala, México and
the United States. The closest relatives of Syrrhophus are the frogs
of the genus Tomodactylus (Dixon, 1957; Firschein, 1954). Lynch
(1968) implied there were no osteological bases for the separation
of Eleutherodactylus, Syrrhophus, and Tomodactylus. At that time,
I believed such to be the case and derived Syrrhophus and Tomodactylus
from the rhodopis complex of Eleutherodactylus, with
which they share terrestrial habits and relatively short limbs. In the
rhodopis complex there is a tendency for the loss of the outer palmar
tubercle, a not uncommon condition in Syrrhophus and Tomodactylus.
However, the skulls of Syrrhophus and Tomodactylus show departures
from the pattern observed in the Middle American Eleutherodactylus,
as well as many of those species in western South
America. Baldauf and Tanzer (1965) reported that the frontoparietals
[Pg 13]
and prootics were fused in Syrrhophus marnockii and that
the prootics and exoccipitals appeared to be one bone (otoccipital).
The otoccipital is not uncommon in eleutherodactyline frogs, but
the fusion of the frontoparietals with the prootics (regardless of the
fusion of the latter with the exoccipital) is uncommon in the family.
I have found the frontoparietal-prootic fusion only in Syrrhophus
(all species), Tomodactylus (all species), and Eleutherodactylus
(West Indies species). None of the Middle American Eleutherodactylus
has the two bones fused. Examination of the character is
difficult in dried skeletal preparations. Cleared and stained or
macerated preparations are satisfactory for checking this character.
Thus, in addition to the presence of numerous plantar supernumerary
tubercles in the frogs of the genera Syrrhophus and Tomodactylus,
these two genera can be separated from other Middle
American eleutherodactylines by the fusion of the frontoparietals
and prootics. This character not only further strengthens the argument
that the two genera are closely related but poses a problem
of zoogeographic analysis of the distribution of the character, which
will be discussed fully elsewhere.
| 1. | Three large, well-developed palmar tubercles | 2 |
| Two large palmar tubercles; outer (third) palmar tubercle reduced in size or absent | 9 | |
| 2. | Digital pads more than twice (usually three or more) times width of digit | 3 |
| Digital pads less than twice width of digit | 4 | |
| 3. | Males having vocal slits; dorsum vermiculate; diameter of tympanum in males about one-half diameter of eye | S. dennisi |
| Males lacking vocal slits; dorsum flecked, spotted, or blotched; diameter of tympanum in male about three-fourths that of eye | S. longipes | |
| 4. | First finger longer than second | 5 |
| First finger shorter than or equal to second | 7 | |
| 5. | Venter smooth; dorsum spotted or vermiculate | S. leprus |
| Venter areolate, or if smooth, dorsum flecked and interorbital bar lacking | 6 | |
| 6. | Venter areolate; interorbital bar present; ground color yellowish | S. cystignathoides cystignathoides |
| Venter smooth; interorbital bar absent; ground color brown | S. cystignathoides campi | |
| 7. | First finger shorter than second; digital tips only slightly dilated; green in life with darker green spots | S. verrucipes |
| First finger equal to second; digital tips slightly to moderately expanded | 8 | |
| 8. | Dorsum vermiculate; interorbital bar present; ground color cream to brown in life [Pg 14] | S. guttilatus |
| Dorsum punctate or flecked; interorbital bar absent; ground color green in life | S. marnockii | |
| 9. | Dorsum dark with pale (red in life) spots; digital pads not expanded | S. rubrimaculatus |
| Dorsum pale with dark markings and digital pads slightly to widely expanded | 10 | |
| 10. | Digital tips not widely expanded; tympanum well-defined; outer metatarsal tubercle more than one-half size of inner | 11 |
| Digital tips widely expanded, truncate in outline; tympanum poorly defined; outer metatarsal tubercle less than one-half size of inner | 12 | |
| 11. | Dorsum dark brown with large light spots or blotches; tympanum/eye ratio usually greater than 43 percent | S. pipilans pipilans |
| Dorsum dark brown with small light spots; tympanum/eye ratio less than 48 percent | S. pipilans nebulosus | |
| 12. | Light interorbital bar present | 13 |
| Light interorbital bar absent | 14 | |
| 13. | Adults small, less than 22 mm. snout-vent length with a broad mid-dorsal stripe; dark bands on shank narrower than light interspaces | S. nivocolimae |
| Adults larger, more than 22 mm. snout-vent length; dorsum vermiculate; dark bands on shank broader than light interspaces | S. interorbitalis | |
| 14. | Dorsum spotted with discrete black spots; pattern definite | S. modestus |
| Dorsum reticulate or vermiculate, pattern poorly defined | 15 | |
| 15. | Adults small, less than 21 mm. snout-vent length; upper arm not banded | S. pallidus |
| Adults larger, usually greater than 21 mm. snout-vent length; upper arm banded | S. teretistes |
The following accounts do not include complete descriptions of each
taxon, because a more than adequate number of descriptions is available
in the recent (1940-1966) literature. An abbreviated synonymy, in which
are listed all combinations and emendations of names and significant
contributions to our knowledge of the taxon, is given for each. For each
species and subspecies the following are given: descriptive diagnosis,
statement of range, remarks on taxonomy, list of specimens examined,
illustration of color pattern, and distribution map.
[Syntypes.—Originally USNM 32402-32409, (32405 now in MCZ) from
Potrero, near Córdoba, Veracruz, México, Francis Sumichrast
collector.]
Diagnosis.—Adults small, males 16.0 to 23.5 mm. in snout-vent length,
females 16.0-25.8 mm. in snout-vent length; vocal slits present in
males; finger tips slightly expanded; first finger longer than second;
outer metatarsal tubercle one-half size of inner, conical, compressed;
skin of dorsum weakly pustular, that of venter smooth to areolate;
tympanum 44 to 69 per cent diameter of eye (mean 55.5 per cent); ground
color yellow to brown in life with brown to black fleckings on dorsum
and flanks; limbs banded; interorbital bar present or not.
Remarks.—Two geographic races (subspecies) are herein recognized;
previously these were held by various authors to be species (campi and
cystignathoides). Intergradation occurs in southern Tamaulipas and
eastern San Luis Potosí, México. The two subspecies can be distinguished
on the basis of color pattern and the condition of the skin of the
venter.
Distribution.—Low to moderate elevations from the Río Grande
embayment to central Veracruz, México (Fig. 5).
Brownsville, Cameron Co., Texas; R. D. Camp collector, March 31, 1915].
Smith and Taylor, 1948:52. Martin, 1958:50.
Diagnosis.—Venter smooth; usually no interorbital light and dark bars
present; ground color brown in life (Fig. 4a).
Remarks.—Martin (1958) was the first author to point out that S.
campi was probably a subspecies of the more southern S.
cystignathoides. Various references in the literature might lead one to
believe that the two were sympatric over much of northeastern México;
this error was created by the use of a single character (condition of
the skin of the venter) to characterize the two populations. Specimens
from southern Texas have a smooth venter, lack interorbital bars and
have, in general, a brown ground color, whereas specimens from central
Veracruz have an areolate venter, interorbital light and dark bars and a
yellow ground color. In southern Tamaulipas and eastern San Luis Potosí,
these characters vary discordantly, thereby strongly suggesting that the
two populations intergrade. Both populations agree in other
morphological characters; therefore, they are here treated as geographic
variants.
Etymology.—Named for the collector of the type specimens, Mr. R. D.
Camp of Brownsville, Texas.
Distribution.—Lower Río Grande embayment in Texas to central Nuevo
León and Tamaulipas, México. Intergrades are known from southern
Tamaulipas and adjacent San Luis Potosí, México (Fig. 5).
Specimens examined.—(113) TEXAS, Cameron Co.: MCZ 10277-85, 10286
(10); Brownsville, AMNH 3215, 3218-20, 3221 (3), 5376, 62117, FMNH
105336, KU 8135-39, MCZ 3738-42, 3743 (10), TCWC 5908, 7139, TNHC 92-94,
20909, UMMZ 51760, 54031 (5), USNM 52290 (holotype); 22 mi. SE
Brownsville, TNMC 14223; 8 mi. SW Brownsville, UMMZ 101127 (3);
Harlingen, AMNH 62118, UMMZ 105200-205, 105206 (5), 105207 (4). Hidalgo
Co.: Bentsen-Río Grande State Park, UMMZ 114378; 6 mi. S McAllen, TNHC
7136-39; Santa Ana Refuge, TCWC 13495-96; Weslaco, TCWC 17658-60.
MEXICO, Nuevo León: Salto Cola de Caballo, AMNH 57953-54, FMNH
30644-45, 37169-70; Monterrey, UIMNH 13324; 40 km. SE Monterrey, UIMNH
3686. Tamaulipas: 80 km. Matamoros, FMNH 27150 (13).
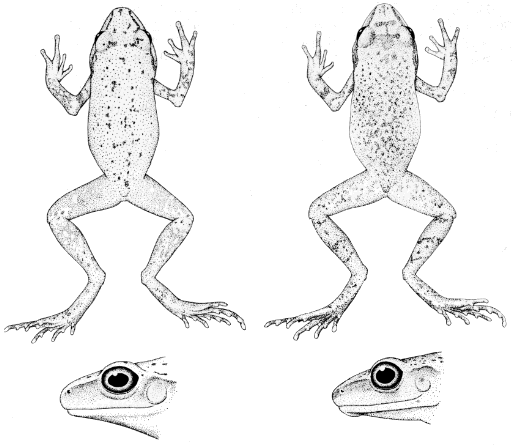
13490) and S. c. cystignathoides (right, KU 105500). Dorsal views ×2,
sides of heads ×3.
Intergrades [S. c. cystignathoides × S. c. campi (88)] MÉXICO, San
Luis Potosí: 5 km. E Ciudad del Maiz, UMMZ 106435; 16 km. W
Naranjo, FMNH 104584; Salto de Agua, 34 km. WSW Antigua Morelos, TCWC
6980. Tamaulipas: 5 km. W Acuña, 1060 m., UMMZ 101172, 101173 (16),
101174-76, 101177 (6); 14.5 km. NNW Chamal, 430 m., UMMZ 111337 (2); 20
km. NNW Chamal, 700 m., UMMZ 111338 (11); 8 km. N Gómez Farías, 450 m.,
UMMZ 101165; 8 km. NE Gómez Farías, Pano Ayuctle, UMMZ 102264, 102924
(6); 8 km. NW Gómez Farías, 1060 m., LSUMZ 11084, UMMZ 101199, 102928
(5), 102929-32, 110124 (3); Río Guayala, near Magiscatzin, MCZ 24138-42,
85071-81, UMMZ 88242 (2); Magiscatzin, TCWC 6981; Las Yucas, north of
Aldama, MCZ 29665-68; 16 km. NE Zamorina, UMMZ 101124.
32402-32409, from Potrero, near Córdoba, Veracruz, México, collected by
Francis Sumichrast]. Boulenger, 1882:196.
Taylor and Smith, 1945: 582-83. Smith and Taylor, 1948:50. Martin,
1958:49.
1946:170].
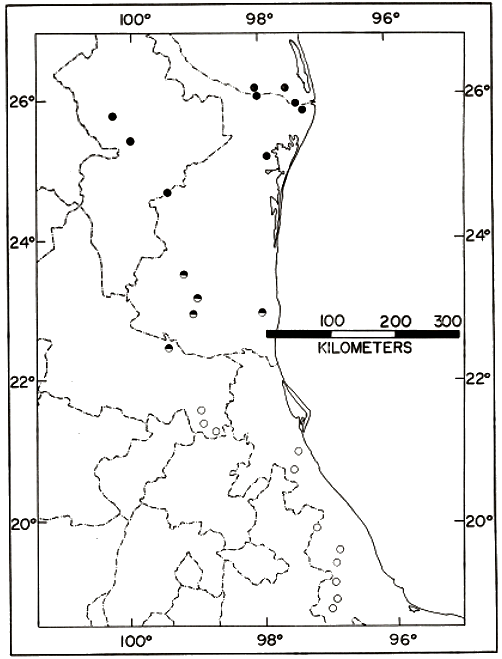
symbols) and the nominate subspecies (open symbols).
Diagnosis.—Venter areolate; interorbital light and dark bars present;
ground color yellow to brownish-yellow in life (Fig. 4b).
Remarks.—Firschein (1954) briefly considered the status of Peters’
(1871) Phyllobates verruculatus and noted that if it was a
Syrrhophus it would probably be referrable to S. cystignathoides.
Peters’ (1871) original description corresponds well with S.
cystignathoides, and the type-locality (“Huanusco” = Huatusco) is
within the range of that species. Firschein (1954) expressed
[Pg 18] doubt that
verruculatus was a Syrrhophus, because Peters placed it in another
genus. However, Peters described verruculatus a decade before Cope
diagnosed the genus Syrrhophus. Most frogs now called Syrrhophus, plus
a number of lower Central American frogs now placed in a variety of
genera were placed in Phyllobates by Boulenger, Cope, and Peters.
The types of Phyllobates verruculatus were destroyed during World War
II (Günther Peters, in litt.); the specimens subsequently assigned to
the taxon by Kellogg (1932) are Syrrhophus cystignathoides. Because
the type specimens are lost and because the name antedates the more
established name, cystignathoides, I favor retaining Phyllobates
verruculatus Peters as a nomen dubium.
Smith and Taylor (1948) reported S. verruculatus from Tianguistengo,
Hidalgo, México. These specimens are examples of verrucipes. Smith
(1947) reported a specimen of verruculatus from San Lorenzo, Veracruz.
Firschein (1954) referred it to cystignathoides, and Duellman (1960)
concluded that both authors were in error and that the specimen (USNM
123530) was a leprus.
Etymology.—The trivial name is the diminutive of Cystignathus, a
once-used generic name for several leptodactylid frogs.
Distribution.—Low and moderate elevations in the foothills along the
Sierra Madre Oriental from eastern San Luis Potosí to Central Veracruz,
México (Fig. 5).
Specimens examined.—(130), MÉXICO, Puebla: Necaxa, UMMZ 69519-20.
San Luis Potosí: 5 km. W Aguismón, LSUMZ 4962-63; along Río Axtla,
road to Xilitla, UMMZ 105500; Tamazunchale, UIMNH 3199; 6.5 km. N
Tamazunchale, UMMZ 104039; 8 km. N Tamazunchale, UMMZ 119490.
Veracruz: Coatepec, 1210 m., FMNH 704966-67; 11 km. SE Coatepec, 850
m., FMNH 70468-70; below Córdoba, FMNH 104588, UIMNH 13321; Cuautlapam,
1000 m., FMNH 106477-80, KU 100364, UIMNH 58200-03, UMMZ 105392; Fortín
de las Flores, UIMNH 13322, 13339; 1.6 km. N Fortín de las Flores, UIMNH
42799-808, UMMZ 105389; 3.2 km. N Fortín de las Flores, UIMNH 26633-35;
4.8 km. N Fortín de las Flores, UIMNH 71967-68; 3.2 km. W Fortín de las
Flores (Barranca Metlac), 910 m., UIMNH 49294-95, UMMZ 115444-46,
118221, 119893 (2); Huatusco, KU 100363; Jalapa, 1400 m., FMNH 70440,
70443-51, 70454-65; 16 km. NE Jalapa, 1300 m., FMNH 70452-53; 8 km. E
Jalapa, UIMNH 13338; 9.5 km. S Jalapa, UMMZ 122083 (2); Mirador, KU
23967; Paraja Nuevo, El Suchil, UMMZ 85490 (7), 85491 (2), 90315; La
Passa, UIMNH 49293, 49297; 1 km. E Plan del Río, 240 m., UMMZ 102067
(2); Potrero Viejo, FMNH 104583, 104586, 105326-27, KU 26789, 100357-62,
UIMNH 13323, 13340-43; USNM 32402 (lectotype), 32403-04, 32406-09; 9.6
km. S Santa Rosa, TCWC 12785; 24 km. NE Tezuitlán (Puebla), UMMZ 105388;
Teocelo, FMNH 70437-38, KU 26080, 26790; 3.2 km. N Teocelo, FMNH 70439,
70441-42; 9.6 km. NW Tihuatlán, UIMNH 3684-85; 15 km. ENE Tlacotepec, KU
23966; 26 km. NW Tuxpan, UMMZ 126419.
Efigena, Oaxaca, México, Francis Sumichrast collector]. Kellogg,
1932:124-5, 128. Taylor and Smith, 1945:582. Smith and Taylor,
1948:50-51. Duellman, 1958:8, pl. 1, Fig. 2; 1960:56-57. Gorham,
1966:165.
Neill collection 1525, from 3.9 mi. N San Antonio, Toledo District,
British Honduras,
collected October 28, 1959, by R. A. Allen, T. C.
Allen, and W. T. Neill].
[Pg 19]Diagnosis.—Medium-sized frogs, males 20.5-26.5 mm. in snout-vent,
females 22.0-29.3 mm. in snout-vent length; vocal slits present in
males; tips of fingers dilated slightly; first finger longer than
second; inner metatarsal tubercle twice size of small, conical outer
metatarsal tubercle; skin of dorsum pustular, that of venter smooth;
snout subacuminate; diameter of tympanum 47.5-62.5 per cent of eye in
males, 38.6-57.9 per cent in females; dorsum yellowish-green with
chocolate brown blotches or spots forming reticulations in most
specimens; venter white to gray; flanks brown, spotted with white or
not; limbs banded; interorbital bar obscured by dorsal pattern.
pattern (left, UMMZ 121244, ×2; right, KU 26106, ×1.7). Side of head
(UIMNH 42726, ×7).
leprus (circles), rubrimaculatus (triangles), and verrucipes
(squares).
Remarks.—My distribution map (Fig. 7) differs somewhat from that of
Duellman (1958), who was unaware of specimens reported by Taylor and
Smith (1945) from central Veracruz, México.
Duellman (1958, 1960) regarded S. leprus as having a gray venter.
Neill (1965) characterized his new subspecies on the basis of white
venter and spots on the dorsum. Some specimens from throughout the range
have only small round spots, instead of vermiculations (Fig. 6). The
gray ventral coloration is largely restricted to the population in Los
Tuxtlas, Veracruz, but only about 80 per cent of the specimens from the
Los Tuxtlas have gray venters, whereas specimens from Guatemala, Oaxaca,
Tabasco, and central Veracruz, México, have white venters (rarely gray).
Since the specimens from British Honduras are not distinct from
specimens throughout most of the range, there is no reason to recognize
them as a subspecies.
Etymology.—Greek, lepra, leprosy, in reference to the mottled color
pattern.
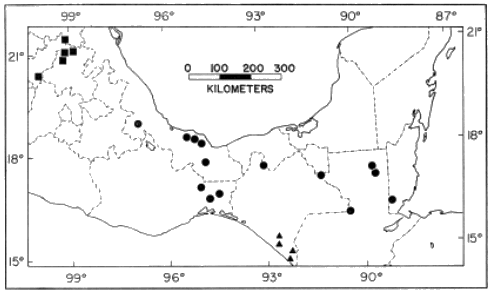
Distribution.—Discontinuous; central Veracruz to British Honduras to
low elevations in the foothills of the Sierra Madre Oriental, Los
Tuxtlas, Sierra Madre de Chiapas (Isthmus of Tehuantepec (Fig. 7)).
Specimens examined.—(84). GUATEMALA, Alta Verapaz: Chinajá, KU
55961-62. El Petén: 15 km. NW Chinajá, KU 55963; Piedras Negras, USNM
114085-92; Tikal, UMMZ 117035; Uaxactún, AMNH 55121-22.
MÉXICO, Oaxaca: Cerro San Pedro del Isthmo, UIMNH 35510; Finca La
Gloria, USNM 114093; 30.5 km. N Matías Romero, UIMNH 39459, 71969; Santa
Efigenia, USNM 10040 (holotype). Tabasco: Teapa, UMMZ 113799-800; 13.5
km. W Teapa, UMMZ 120253. Veracruz: 27.5 km. N Acayucan, UIMNH 42726;
Atoyac, UIMNH 13331, 49296; 3.2 km. N Catemaco, UIMNH 71976-77; Coyame,
UIMNH 38995, 38998, 40342; Dos Amates, TCWC 21211; Fortín de Las Flores,
FMNH 113751, 113753; Paraja Nuevo, El Suchil, UMMZ 90315; Potrero Viejo,
FMNH 113743-50, 126114-18, KU 26104-06, UIMNH 13332-37, UMMZ 88837; San
Andrés Tuxtla, UIMNH 27123-31, 28611, 71975, UMMZ 115450 (5); San
Lorenzo, USNM 123530; 4.5 km. NW Santiago Tuxtla, JDL 992 (skeleton),
UIMNH 27122; 32 km. S Sayula, EAL 1696; Tepalapan, 1.6
[Pg 21]
km. S Catemaco, UMMZ 118222 (2); Volcán San Martín, south slope,
UMMZ 118223; Volcán San Martín, Rancho El Tular, UIMNH 35399-400, 40340-41.
[Holotype.—USNM 114070, from La Esperanza, near Escuintla, Chiapas,
México, collected May 13, 1940, by H. M. and R. Smith]. Duellman,
1958:1-4, 7, 12, 14. Gorham, 1966:167.
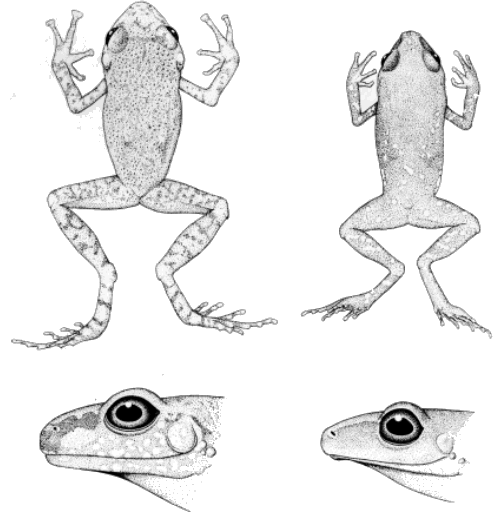
right, KU 58910, ×4) and S. verrucipes (upper left, UIMNH 15995, ×1.6;
lower left, UIMNH 15989, ×3.7).
Diagnosis.—Small frogs, males 18.2-23.5 mm. snout-vent, females
19.0-22.5 mm. snout-vent length (small sample); vocal slits in males;
digital tips scarcely expanded (Fig. 1); first finger shorter than
second; outer palmar tubercle reduced in size; inner metatarsal tubercle
elongate, twice the size of small, conical outer metatarsal tubercle;
diameter of tympanum 35.5-46.5 per cent that of
[Pg 22]
eye in both sexes; dorsum brown with small pale spots (red in life); venter gray.
Remarks.—Previous authors who treated Syrrhophus placed this
species in the western complex, because it occurs on the Pacific versant
and has a reduced outer palmar tubercle. Duellman (1958) placed
rubrimaculatus apart from the other western species, because of its
relatively unexpanded digital tips and coloration. The digital tips are
like those in leprus, which
rubrimaculatus
resembles. Except for the reduction of the outer palmar tubercle,
rubrimaculatus could be a member of the leprus group.
Syrrhophus rubrimaculatus is probably best treated as a Pacific
derivative of the leprus group, even though the palmar tubercles do
not agree. The removal of rubrimaculatus from the western complex
results in a more homogeneous remainder and does not greatly increase
the heterogeneity of the eastern complex.
Etymology.—Latin, meaning spotted with red; in reference to the
colors in life.
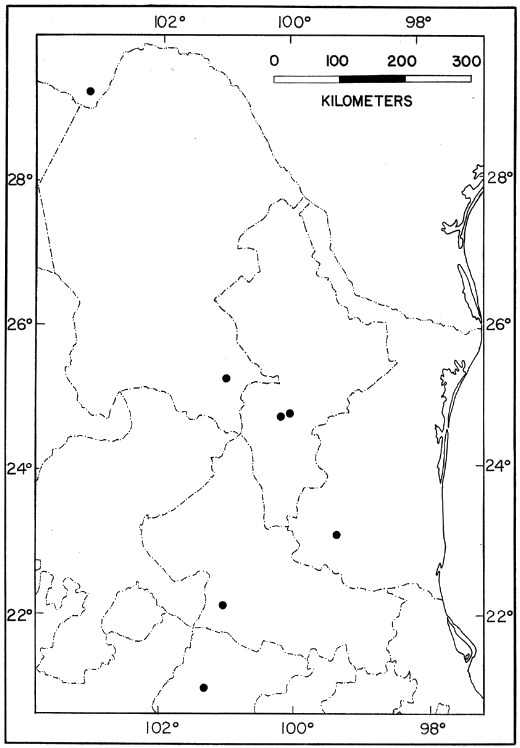
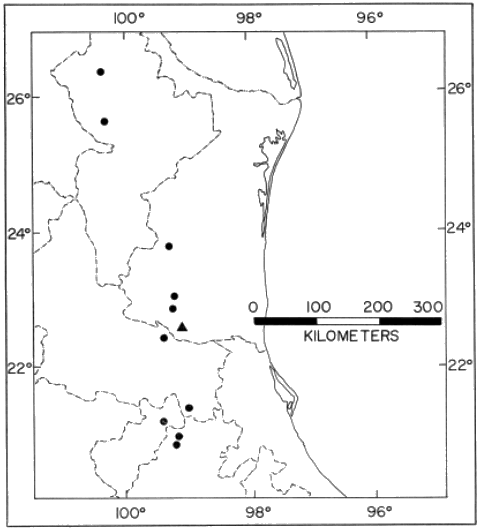
Distribution.—Low to moderate elevations on the Pacific versant of
southeastern Chiapas, México (Fig. 7); probably extending into
adjacent Guatemala.
Specimens examined.—(48) MÉXICO, Chiapas: Escuintla, UMMZ 88283; 6
km. NE Escuintla, UMMZ 87876-80; La Esperanza, UIMNH 13285, UMMZ
88496-97, USNM 114070 (holotype), 114054-69, 114072; Monte Cristo, UMMZ
88353; 1.3 km. N Puerto Madero, KU 58910-11; Finca San Jerónimo, 600-650
m., UIMNH 55299-312, 55313-16 (cleared and stained).
Guanajuato, Guanajuato, México; collected in 1877 by Alfredo Duges].
127-28. Smith and Taylor, 1948:49, 51. Firschein, 1954:52-54. Gorham,
1966:164.
from 15 mi. SW Galeana, Nuevo León, México, 1575 m.; collected on
October 13, 1939, by Hobart M. Smith]. Smith and Taylor, 1948:49, 51.
Firschein, 1954:54-55. Martin, 1958:50. Gorham, 1966:167.
from the Basin, Chisos Mountains, Brewster Co., Texas; collected on July
24, 1937, by Walter L. Necker].
from 5 km. SW San Luis Potosí, San Luis Potosí, México; collected on
July 18, 1949, by David Langebartel]. Gorham, 1966:166.
part).
Diagnosis.—Medium-sized frogs, males 20.6-29.0 mm. snout-vent,
females 25.7-31.0 mm. snout-vent length; vocal slits in males; digital
tips slightly expanded (Fig. 1); first and second fingers equal; skin of
dorsum smooth to moderately pustular, that of venter smooth; snout
blunt; diameter of tympanum 55.1-75.7 per cent that of eye in males,
47.6-61.7 in females; dorsum and flanks cream to gray with light brown
to black flecking and vermiculations; thighs usually not banded;
interorbital bar present (Fig. 8).
left, UIMNH 55519, ×2.3) and S. marnockii (upper right, TCWC 9317,
×1.4; lower right, TCWC 13510, ×2.1).
Remarks.—Cope (1879) distinguished Malachylodes from Syrrhophus
on the basis of the presence of a frontoparietal fontanelle in the
holotype of guttilatus. The holotype is a juvenile female and as is
the case in the juveniles of nearly all leptodactylids, a frontoparietal
fontanelle is present. Firschein (1954) used the presence of the
fontanelle to distinguish guttilatus from his petrophilus.
As is clearly evident from the length of the synonymy, I consider a
number of currently used names to be synonymous with guttilatus. I
have seen the holotypes of all four names and am unable to recognize
more than a single species. The holotype of petrophilus is a male,
whereas that of smithi is a female. The supposed differences are a
reflection of sexual dimorphism in the size of the eye (Table 5). The
two holotypes, as well as those of gaigeae and Malachylodes
guttilatus agree in color pattern.
Schmidt and Smith (1944) named Syrrhophus gaigeae from the Chisos
Mountains of the Big Bend region of Texas and compared it only with S.
marnockii. Milstead, Mecham and McClintock (1950) synonymized gaigeae
and marnockii because they were unable to verify the characters Wright
and Wright (1949) used to separate them. Specimens from the Big Bend
region differ from those of the Edward and Stockton Plateaus in having a vermiculate
[Pg 24]
pattern, an interorbital bar, and a supratympanic stripe.
In these respects they agree with specimens from northern México. Based
on limited observations, the Mexican population is yellowish to brownish
in life whereas the central Texas population is green in life. Lacking
evidence of genetic exchange, the two are held to be specifically
distinct.
[Pg 25]
Nearly every specimen examined was infested with chiggers of the genus
Hannemania. The greatest concentrations are on the venter, in the
groin, and on the thighs. Many specimens have chiggers on the digits and
tarsi. The same, or a related, chigger was found on many specimens of
Syrrhophus marnockii and a few S. verrucipes, but on no other
species of the genus. Mr. Willy Wrenn told me that he has seen heavy
infestations of Hannemania on Syrrhophus pallidus. Infestation by
Hannemania probably reflects similar ecologies rather than close
relationships.
Etymology.—Latin, guttula, meaning spotting or flecking, in
reference to the color pattern.
Distribution.—Moderate to intermediate elevations (600 to 2000 m.)
along the Sierra Madre Oriental from the Big Bend Region of Texas to
Guanajuato, México (Fig. 10).
Specimens examined.—(32) TEXAS, Brewster Co.: Juniper Canyon,
Chisos Mts., FMNH 27361 (holotype of S. gaigeae), 27360, 27362-63, MCZ
15346, 27801, UMMZ 66080, 66082, 66085-91, USNM 76876; Upper Green
Gulch, TCWC 15943.
MÉXICO: Coahuila: 8 km. S Saltillo, UIMNH 55518-21. Guanajuato:
Guanajuato, USNM 9888 (holotype of Malachulodes guttilatus); 8 km. E
Guanajuato, AMNH 73425; Cerro Cubilete, AMNH 73424. Nuevo León: 3 km.
S Galeana, JDL 1215 (skeleton), UIMNH 58204; 24 km. SW Galeana. 1575 m.,
USNM 108594 (holotype of Syrrhophus smithi). San Luis Potosí: 5 km.
SW San Luis Potosí, UIMNH 7807 (holotype of S. petrophilus).
Tamaulipas: 1.6 km. NW La Joya de Salas, 1530 m., UMMZ 110736 (4).
“near San Antonio,” Bexar Co., Texas; collected by G. W. Marnock].
McClintock, 1950:550.
Diagnosis.—Medium-sized frogs, males 18.4-28.9 mm. snout-vent,
females 20.4-35.4 mm. snout-vent length; vocal slits in males; digital
tips widened (Fig. 1); first and second fingers equal; skin of dorsum
smooth to weakly pustular, that of venter smooth; snout blunt, rounded;
diameter of tympanum 47.2-68.3 per cent that of eye in males, 45.8-73.3
in females; dorsum tan to light brown in preservative with rusty-brown
flecks, venter white; ground color green in life; thighs banded;
interorbital bar absent.
Remarks.—Specimens from the southern edge of the Edwards Plateau and
the eastern edge of the Stockton Plateau have larger flecks on the back
that tend to form a vermiculate pattern like that of S. guttilatus.
The vermiculation is never well developed (see plate 38 in Conant,
1958). Most of the specimens from the Edwards Plateau have a punctate
pattern (Fig. 9).
Fossils are known from the Sangamon interglacial deposits in Foard and
Knox Counties, Texas (Lynch, 1964; Tihen, 1960).
Etymology.—A patronym for the collector of the type specimens.
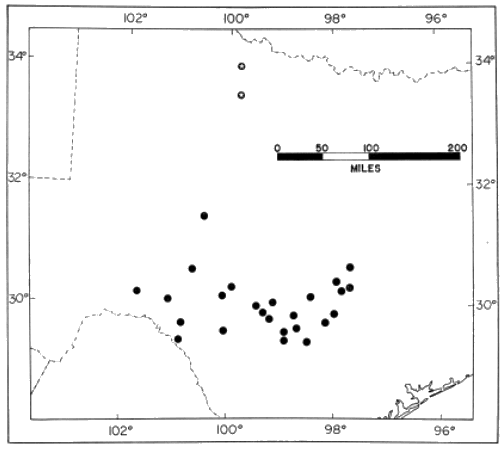
Distribution.—The Edwards Plateau and the extreme eastern edge of the
Stockton Plateau in Texas (Fig. 11). The fossil records lie some 200
miles to the north. Two specimens (FMNH 103216-17) from Brownsville,
Cameron Co., Texas, were formerly in the EHT-HMS collection (nos.
31348-49). Data given in Taylor’s field catalogue (housed in the
Division of Reptiles, Field Museum) are “Brownsville, A. J. Kirn
collector, April 15, 1934.” Until verification
[Pg 26]
by recently collected material is available, this record must be disregarded.
Specimens examined.—(103) TEXAS, Bandera Co.: 10 mi. SW Medina,
TCWC 13508-10; 8 mi. W Medina, KU 60243; 13 mi. W Medina, KU 60242, TCWC
13506-07. Bexar Co.: UIMNH 34694; Classen ranch, near San Antonio.
UMMZ 98891; Helotes, EAL 1560, MCZ 11837 (2), UMMZ 64045, USNM 13635; 2
mi. N Helotes, TCWC 9234-35; 3.5 mi. N Helotes, LSUMZ 10363; 8 mi. N
Helotes, TCWC 1549, 4364; San Antonio, FMNH 15553-56, TCWC 13497-99.
Blanco Co.: 8 mi. NE Blanco, TCWC 4782. Comal Co.: New Braunfels,
TCWC 13500-05; 5 mi. NE New Braunfels, UMMZ 71016 (10). Hays Co.: San
Marcos, AMNH 22661-64, 32700, FMNH 15245-46, 26250, 26253-57, 37617,
37665, MCZ 15649-50, 23268-69; 6 mi. SW San Marcos, TCWC 5070-71, 7140,
9232-33, 9236, 9316-17, 9320. Kendall Co.: 11 mi. E Boerne, AMNH
54660-61, 54662 (2); 10 mi. W Boerne, KU 18441; Kendalia, UIMNH 21434.
Kerr Co.: Kerr W. M. Area, TCWC 15859; 40 mi. NW Kerrville, TCWC 6555.
Medina Co.: UIMNH 13287-88; 12 mi. N Castroville, UIMNH 21423; 14 mi.
N Castroville, UIMNH 21424-25; 16 mi. N Castroville, UIMNH 21421-22; 17
mi. N Castroville, UIMNH 21428-29; 18 mi. N Castroville, UIMNH 21426-27,
21430-33; 6.5 mi. NW Rio Medina, KU 18440. Real Co.: Rio Frio, FMNH
55156-57. Travis Co.: Austin, AMNH 44221-22; Mount Bonnell, 5 mi. S
Austin, UMMZ 101453 (10). Uvalde Co.: 13 mi. from Uvalde, UIMNH 62322.
Val-Verde Co.: 40 mi. N Del Rio, JDL 214 (skeleton).
localities are late Pleistocene records.
Zacualtipán, Hidalgo, México (1800 feet lower in a rocky gorge of a
stream near its junction with the Río San Miguel), collected by Dr.
Santiago Bernard]. Kellogg, 1932:126-29. Smith and Taylor, 1948:52-53.
Firschein, 1954:55-57. Gorham, 1966:167.
[Holotype.—FMNH 100049 (formerly EHT-HMS 6838), from La Placita, 8 km.
S Jacala, Hidalgo, México, 1850 m.; collected on July 2, 1936, by Edward
H. Taylor]. Smith and Taylor, 1948:47-48.
Diagnosis.—Medium-sized frogs, males 17.5-26.1 mm. snout-vent,
females 28.0-31.7 mm. snout-vent length; vocal slits in males; digital
tips slightly expanded; first finger shorter than second; skin of dorsum
pustular, that of venter areolate; snout elongate, subacuminate;
diameter of tympanum 56.1-76.7 per cent that of eye in males, 54.3-56.8
in females; in preservative, dorsum reddish brown with numerous small
black or dark brown spots (Fig. 8); venter white to cream; in life
dorsum green with darker green spots, belly white; iris gold above,
bronze below.
Remarks.—Cope’s (1885) original description was not sufficiently
clear to enable subsequent authors to recognize this species. Taylor
(1940e) described it as a Tomodactylus, but Dixon (1957) pointed out
that T. macrotympanum differed from the other species of the genus in
having a poorly developed lumbo-inguinal (inguinal) gland, and placed
the species in the genus Syrrhophus. Comparison of the holotypes of
S. verrucipes and T. macrotympanum leaves no doubt in my mind that a
single species is involved. This same species was reported by Smith and
Taylor (1948) as S. verruculatus.
Syrrhophus verrucipes bears
resemblance to members of
both the leprus and marnockii groups. In snout shape it is closer to
the leprus group, whereas in digital pad, the shape of the general
body form, and contiguity of habitat it is most similar to the
marnockii group (S. guttilatus).
Etymology.—Latin, meaning warty foot, probably in reference to the
numerous plantar supernumerary tubercles.
Distribution.—Moderate elevations in southeastern San Luis Potosí,
Queretaro, and northwestern Hidalgo, México (Fig. 7).
Specimens examined—(43) MÉXICO, Hidalgo: Jacala, UMMZ 106434; 9.6
km. NE Jacala, Puerto de la Zorra, 1820 m., KU 60240-41, TCWC 11090,
11147; 8 km. S Jacala, La Placita, 1850 m., FMNH 100049 (holotype of
Tomodactylus macrotympanum), 100791-803, 105334-35, 114287, UIMNH
15989-92, 15995-96, UMMZ 117252, USNM 137202; Tianguistengo, FMNH
113705-09, UIMNH 13328-30; near Zacualtipán, ANSP 11325 (holotype of
Syrrhophus verrucipes). Queretaro: 3.5 km. S San Juan del Río, EAL
1343. San Luis Potosí: 9.6 km. W Ahuacatlán, LSUMZ 4968-70.
Holotype.—UMMZ 101121, adult male from a cave near El Pachón, 8 km. N
Antiguo Morelos, Tamaulipas, México, 250 m., collected on March 13,
1949, by Paul S. Martin.
[Pg 28]
Paratopotypes.—(26). UMMZ 101122 (10), 101123 (2), 101126, 126993
(12).
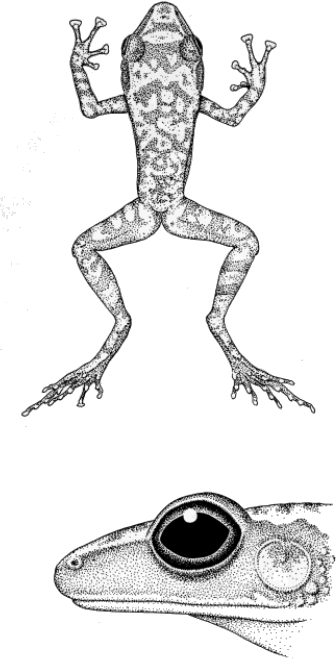
Diagnosis.—Medium-sized frogs, males 22.8-28.4 mm. snout-vent,
females 25.9-32.0 mm. snout-vent; vocal slits in males; digital tips
greatly expanded, more than twice width of digit; first finger shorter
than second; skin of dorsum shagreened to pustular, that of venter
weakly to moderately areolate; toes webbed basally; dorsum light brown
to tan with brown vermiculations; venter white; diameter of tympanum
53.9 to 64.2 per cent that of eye in males, 50.6 to 58.7 per cent in
females.
Description and variation.—(Fig. 12). Head wider than body; head as
wide or wider than long in males, sometimes longer than wide in females;
snout acuminate in dorsal view, elongate and rounded in lateral profile;
canthus rostralis rounded but distinct; loreal region slightly concave,
sloping abruptly to lip; lips not flared; eyelid about two-thirds
interorbital distance; length of eye less than distance between eye and
nostril; diameter of tympanum 53.9 to 64.2 per cent that of eye in
males, 50.6 to 58.7 per cent in females; tympanum round and distinct in
both sexes; supratympanic fold moderately distinct; choanae within
border of jaws, completely visible from directly below, rounded to
slightly oval; dentigerous processes of prevomers and teeth absent;
tongue free for posterior one-half, generally oval in outline; vocal
slits present in males.
Many scattered pustules on dorsum; flanks areolate; skin of venter
areolate or not (variability may be due to differences in preservation);
ventral disc distinct on chest and lower abdomen; inguinal gland present
or not, when present varying from very large and distinct to poorly
defined; axillary gland absent.
First finger shorter than second; all fingers bearing truncate tips with
pads, each pad having a terminal groove; fingers fringed; fingers three
and four having dilated pads two to three times width of digit;
subarticular tubercles large, conical, rounded, simple; supernumerary
tubercles numerous on thenar surface, none on digits; three palmar
tubercles, outer slightly smaller than largest supernumerary tubercles;
row of tubercles on outer edge of forearm variable, weak to very
distinct; tips of toes wider than digits, rounded to truncate at tips,
each pad having terminal groove; toes having lateral fringes, bases of
toes united by web, web not extending to basal subarticular tubercle;
subarticular tubercles smaller than those of hand, round, conical,
simple; supernumerary tubercles numerous on plantar surfaces, extending
between metatarsal tubercles, present on toes between basal two
subarticular tubercles in some specimens; outer metatarsal tubercle
round, conical, one-half as large as ovoid, non-compressed inner
metatarsal tubercle; tarsal tubercles or folds absent.
Ground color pale reddish-brown to tan dorsally, creamy on flanks;
dorsal pattern consisting of reddish-brown to brown vermiculations
extending onto flanks; distinct interorbital light bar present; loreal
region darker than snout, reddish-brown compared to tan or pale
reddish-brown; arms colored like dorsum; thighs banded, unicolor brown
on posterior surfaces; shanks and tarsi banded; venter white to cream
punctated with brown in some specimens.
The variation in proportions is summarized in Table 5.
Remarks.—Martin (1958) expressed some doubt that this series of
26 specimens was identical with “S. latodactylus.” My study indicates
that the specimens from El Pachón represent a distinctive but allied
species. Males of the two species can be readily separated by the
relative sizes of the tympani, presence or absence of vocal slits, and
color pattern. Females of the two species can be separated by color
pattern. Within the type-series, the pattern varies from weakly to
strongly vermiculate but is always recognizable as vermiculate rather
than spotted as in S. longipes (= S. latodactylus of Taylor and
Martin).
[Pg 30]
Etymology.—The specific name is a patronym for David M. Dennis, whose
drawings greatly enhance the worth of this paper.
Distribution.—Known only from the type series.
[Holotype.—apparently USNM 3237 (cited as 3207 by Cope, 1887:16), now
lost, from 40 Leagues from (probably north) México City; collected by
John Potts]. Kellogg, 1932:107.
Taylor, 1948:61. Lynch, 1963:580-581. Gorham, 1966:82.
fig. 7 [Holotype.—FMNH 100063 (formerly EHT-HMS 6807), from Huasteca
Canyon, 15 km. W Monterrey, Nuevo León, México, 680 m.; collected on
June 20, 1936, by Edward H. Taylor]. Smith and Taylor, 1948:50-52.
Martin, 1958:48-50. Gorham, 1966:165.
Diagnosis.—Large frogs, males 22.1-33.2 mm. snout-vent, females
26.8-39.6 mm. snout-vent length; vocal slits lacking in males; digital
tips greatly expanded (more than twice the width of digit); first finger
shorter than second; skin of dorsum pustular, that of venter smooth;
diameter of tympanum in males 61.1-87.2 per cent that of eye, 49.5-72.1
per cent in females; dorsum tan with large or small spots and blotches;
limbs banded; interorbital bar or triangle present.
Remarks.—I have applied Baird’s Batrachyla longipes to the frog
Taylor (1940d) called Syrrhophus latodactylus because the color
pattern (Fig. 13) predominant in the southern part of the range agrees
with that described (figured) for Batrachyla longipes.
The color pattern of individuals in the southern part of the range of
this species consists of large spots or blotches, whereas in the
northwestern part the pattern is made up of smaller spots. In the
northeastern part of the range, the pattern is more reduced and tends to
consist of heavy flecking. The interorbital bar is narrower in specimens
from Nuevo León and Tamaulipas and is triangular in specimens from
Hidalgo and Queretaro.
The status of the name Batrachyla longipes is currently that of a
nomen dubium (Lynch, 1963). At that time, I was unaware of the
geographic variation in color pattern in Syrrhophus latodactylus.
The exact type-locality of Batrachyla longipes is not known. If it is
40 Leagues north of México City, the locality would be in an area where
the species has a blotched instead of a flecked or spotted pattern. No
justifiable evidence was presented to place Batrachyla longipes in
Eleutherodactylus instead of Syrrhophus. Barbour (1923) and Kellogg
(1932) associated another species (E. batrachylus) with longipes.
Taylor (1940a) noted this as a case of misidentification and corrected
the error but left longipes in the genus Eleutherodactylus. Lynch
(1963) noted several points of morphological agreement between
Syrrhophus and B. longipes but did not place longipes in
Syrrhophus.
Baird’s (1859) figures of the holotype do not illustrate prevomerine
teeth, but according to Cope (1866) they were present in the holotype.
The digital
[Pg 31]
tips of the frog in the figure are somewhat narrower than
those typically seen in S. latodactylus. If the specimen was slightly
desiccated, as possibly was the case, the digits would appear narrower.
There is no evidence contrary to placing Syrrhophus latodactylus in
the synonymy of Batrachyla longipes.
variation in pattern (left, TCWC 12179, ×1.5; right, KU 92572, ×1.8);
side of head (TCWC 10966, ×6).
Application of Baird’s name Batrachyla longipes to the species of frog
heretofore called Syrrhophus latodactylus poses one serious problem. Batrachyla
[Pg 32]
longipes is the type-species (by original designation) of the genus Epirhexis
Cope, 1866, which has priority over Syrrhophus Cope, 1878. If Batrachyla
longipes is left in the status of a nomen dubium, Epirhexis can be forgotten,
for the two names are tied together. However, since it seems almost certain
that Batrachyla longipes and Syrrhophus latodactylus are conspecific, the former
name should not be left as a nomen dubium. Epirhexis never came into general
usage (Cope cited the name four times, but no one else has used it), whereas
Syrrhophus is well established in the zoological literature. It would serve only
to confuse the literature to adhere strictly to the Law of Priority and replace
Syrrhophus with Epirhexis. Therefore, Syrrhophus is used in this paper, even
though Epirhexis has priority. A request for the suppression of Epirhexis Cope,
1866, has been submitted to the International Commission of Zoological
Nomenclature (Lynch, 1967).
Etymology.—Latin, meaning long-footed; Taylor’s latodactylus refers to
the wide digital pads.
[Pg 33]
Distribution.—Moderate elevations (650 to 2000 meters) along the
Sierra Madre Oriental from central Nuevo León to northern Hidalgo,
México (Fig. 14).
Specimens examined.—(122) MÉXICO, Hidalgo: 3 km. NE Jacala, AMNH
52977; 9.6 km. NE Jacala, 1800 m., TCWC 10966-70, 12179; 8 km. S Jacala,
La Placita, 1850 m., FMNH 100266-68, 103244, UIMNH 13291, 13327. Nuevo
León: Salto Cola de Caballo, KU 92572; Huasteca Canyon, 15 km. W
Monterrey, 680 m., FMNH 100063 (holotype of S. latodactylus), UIMNH
13290; 6.5 km. N Pablillo, EAL 1319; Sabinas Hidalgo, USNM 139728.
Queretaro: Cueva de los Riscos, 8 km. SW Jalpan, KU 106300. San Luis
Potosí: 13 km. E Santa Barberita, LSUMZ 2295; second camp, San Luis
Potosí road, UIMNH 13326; Xilitla, Cueva sin nombre, UMMZ 125892.
Tamaulipas: 4 km. W El Carrizo, 500 m., UMMZ 111343 (31); 8 km. N
Chamal, Bee Cave, KU 106299; 14.5 km. NNW Chamal, 420 m., UMMZ
111339-40, 111342 (4), 111344 (11); 19 km. NNW Chamal, 700 m., UMMZ
111341 (3); El Chihue, 1880 m., UMMZ 111289 (4); 11 km. N Gómez Farías,
1060 m., UMMZ 101166; 11 km. WNW Gómez Farías, 1800 m., UMMZ 108507 (3);
8 km. NW Gómez Farías, 1060-1400 m., LSUMZ 11085, UMMZ 101167 (3),
101168 (4), 101169 (2), 101170 (3), 101171 (2), 101360-61, 102860,
102933 (4), 102934 (2), 102935-38, 102939 (2), 102940-43, 108800 (3),
110735, 111345-46.
(formerly EHT-HMS 6843), 14.6 km. S Mazatlán, Guerrero, México;
collected on July 22, 1936, by Edward H. Taylor].
Diagnosis.—Medium sized frogs, males 22.6-28.5 mm. snout-vent,
females 21.1-29.4 mm. snout-vent length; vocal slits present in males;
finger tips slightly expanded, truncate in outline; inner metatarsal
tubercle less than twice the size of outer; skin of dorsum smooth to
shagreened, that of venter smooth; tympanum 36.5-54.0 per cent diameter
of eye; dorsum dark brown with large or small light brown, orange-brown,
or yellowish spots or blotches; limbs banded; interorbital bar absent.
subspecies of Syrrhophus pipilans. N = 17 in nebulosus, 18 in
pipilans.
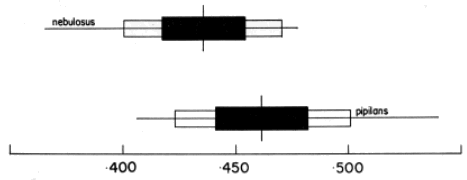
Remarks.—Two subspecies were recognized by Duellman (1958).
Previously both had been treated as species. The two populations were
distinguished on the basis of color pattern and the size of the
tympanum. Measurements of 17 males of S. p. nebulosus from central
Chiapas and 18 males of S. p. pipilans from south-central Oaxaca and
Guerrero, México, demonstrates that the supposed difference in tympanum
size is not significant (Fig. 15).
[Pg 34]
There is, however, a tendency for
the western population of S. pipilans to have larger tympani. Based on
the present examination of 112 specimens of this species the two
populations are held to be sufficiently distinct to warrant taxonomic
recognition as subspecies (Fig. 16).

The parotoid glands attributed to this species by Taylor (1940c:95) are
merely the superficial expression of the m. depressor mandibulae and
scapula. No true glands are present in the parotoid region.
[Holotype.—FMNH 100095 (formerly EHT-HMS 3774), near Tonolá, Chiapas,
México; collected on August 27, 1935, by Hobart M. Smith and Edward H.
Taylor]. Smith and Taylor, 1948:49, 51.
1963:32-33. Gorham, 1966:166-67.
Diagnosis.—Diameter of tympanum 36.6-47.8 per cent that of eye;
dorsum dark brown with numerous small light brown to yellowish spots.
Remarks.—The distribution of this subspecies is adequately described
by Duellman (1958). Fouquette (1960) described the vocalization of this
frog.
Etymology.—Latin, nebula, in reference to the clouded dorsal
pattern.
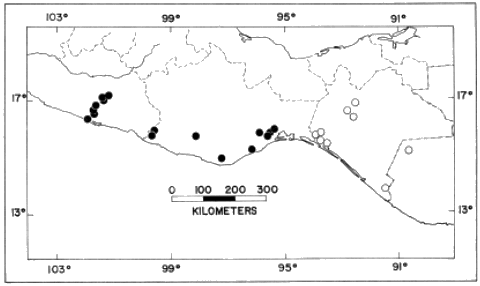
Distribution.—Low to moderate elevations along the Pacific versant of
Chiapas and in the Grijalva valley of Chiapas and Guatemala (Fig. 17).
Specimens examined.—(54) GUATEMALA, Huehuetenango: Jacaltenango,
UMMZ 117036; 35 km. SE La Mesilla, TNHC 29652. MÉXICO, Chiapas: 11.2
km. N Arriaga, 300 m., UMMZ 125891; 11.8 km. N Arriaga, UMMZ 117279;
[Pg 35]
12.8 km. N Arriaga, UMMZ 117280; 17.5 km. S Arriaga, UIMNH 57108-109;
1.5 km. S Bochil, 1250 m., KU 58898-908; Cerro Hueco, 7 km. S Tuxtla
Gutierrez, UMMZ 123007; 3.2 km. S Ixtapa, UMMZ 124000; Linda Vista, ca.
2 km. NW Pueblo Nuevo Solistahuacán, KU 58897; Hda. Monserrate, 40 km.
NW Arriaga, UMMZ 102258; near San Ricardo, FMNH 100720; Tapachula, FMNH
75792, 103242, 100695-96, UIMNH 13292; 56 km. E Tapanatepec, Oaxaca,
TNHC 26942, Tonolá, FMNH 100095 (holotype), 100686-92, UIMNH 13293-95;
Tuxtla Gutierrez, FMNH 100693-94, UIMNH 13297; 19 km. N Tuxtla
Gutierrez, TNHC 25229-30; 15.5 km. NE Tuxtla Gutierrez, UMMZ 119892 (3);
19 km. NE Tuxtla Gutierrez, UMMZ 119891 (3); 8 km. NNW Tuxtla Gutierrez,
KU 37809; Unión de Juarez, FMNH 105294.
(formerly EHT-HMS 6843), from 14.6 km. S Mazatlán, Guerrero, México;
collected on July 22, 1936, by Edward H. Taylor]. Taylor and Smith,
1945:581-82. Smith and Taylor, 1948:49, 50-51.
fig. 1. Gorham, 1966:166.
Diagnosis.—Diameter of tympanum 40.6-54.0 per cent that of eye;
dorsum dark brown with large light spots or blotches.
Remarks.—Duellman’s (1958) synopsis of this subspecies is adequate;
the distribution has not been extended, but several records are now
available which fill in gaps.
circles) and pipilans (solid circles).
Gadow’s (1905) record of S. verruculatus from “Buena Vista, S.
Guerrero” is most likely applicable to this species. Gadow simply
included the name in a list of the species he had collected during his
trip in México (1902-04); no
[Pg 36]
further comment was made on this species
although references to Syrrhopus (sic) appear in several places in the
paper and would appear to apply to the species he had.
Etymology.—Latin, pipilo, chirping, peeping, in reference to the
call of the male.
Distribution.—Sea level to about 1800 meters along the Pacific
versant of western México from central Guerrero to the Isthmus of
Tehuantepec (Fig. 17).
Specimens examined.—(62). MÉXICO, Guerrero: Acapulco, UMMZ 110125;
6.4 km. N Acapulco, FMNH 100389, 100525; Agua del Obispo, 980-1000 m.,
FMNH 75791, 100518-21, 100526, KU 86884-86, UIMNH 13315, UMMZ 119152,
125890 (4); 13.3 km. NW Coyuca, UIMNH 38367, 71982-83; 14.5 km. S
Mazatlán, FMNH 100072 (holotype), 100408, 100511-17, UIMNH 13302-309;
Tierra Colorado, 300 m., KU 67961, UIMNH 13313-14; near El Treinte, FMNH
126639; Xaltinanguis, FMNH 100522-24, 126640. Oaxaca: Cacahuatepec,
UIMNH 52853; 8 km. NW Río Canoa, 53 km. ESE Cuajinicuilapa, UIMNH
52852; 6.4 km. N El Candelaria, UIMNH 9501; 11.2 km. S El Candelaria,
UIMNH 9502; 17 km. NE Juchatengo, 1600 m., KU 86887; 31.5 km. N
Pochutla, UMMZ 123999 (2); 32.9 km. N Pochutla, 850 m., UMMZ 123996;
37.1 km. N Pochutla, UMMZ 123998 (2); 41.4 km. N Pochutla, UMMZ 123997
(2); Cerro Quiengola, FMNH 105653; 3.8 km. N Santiago Chivela, UMMZ
115449; 14.5 km. W Tehuantepec, UMMZ 115448 (2).
1-2 [Holotype.—UIMNH 67061 (formerly FAS 9378), 36 mi. N Mazatlán,
Sinaloa, México, collected on November 17, 1955, by E. C. Bay, J. C.
Schaffner, and D. A. Langebartel]. Duellman, 1958:1-4, 10, 12, 14.
Gorham, 1966:164-65.
S. nivocolimae (LACM 3203, ×1.3), and S. teretistes (KU 75263,
×1.5).
Diagnosis.—Medium sized frogs, only known male 25.6 mm. snout-vent,
females 20.0-26.7 mm. snout-vent length (small sample); vocal slits in males;
[Pg 37]
finger tips expanded; first finger shorter than second; outer
metatarsal tubercle one-third size of inner; skin of dorsum shagreened,
that of venter smooth; diameter of tympanum 37.7-42.4 per cent that of
eye in both sexes; pale yellow-brown ground color mottled with brown;
limb bands broad, much wider than narrow light interspaces; interorbital
bar very long, edged with dark brown to black (Fig. 18).
Remarks.—Duellman’s (1958) measurements and proportions of S.
interorbitalis were based exclusively on the type series, which is
composed of only females; therefore his interorbitalis data are not
comparable with the data for the other species in his table. Campbell
and Simmons (1962) collected the only known male. The type series was
collected beneath rocks in a stream bed; the collectors heard calling
frogs in the bushes but were unable to obtain specimens (Langebartel and
Shannon, 1956). Campbell and Simmons (1962) reported that their specimen
had a poorly developed interorbital bar in life; in preservative the bar
compares favorably with the bar in the female (Fig. 18).
Etymology.—Latin, in reference to the pale interocular band.
Distribution.—Pacific lowlands of Sinaloa, México (Fig. 20).
Specimens examined.—(10). MÉXICO, Sinaloa: 36 mi. N Mazatlán, UIMNH
38094-96, 67061 (holotype), 71970-74; 65 mi. N Mazatlán, LACM 13773.
100048 (formerly EHT-HMS 3756), from Hacienda Paso del Río, Colima,
México; collected on July 8, 1935, by Hobart M. Smith]. Smith and
Taylor, 1948:49-50.
1. Gorham, 1966:166.
Diagnosis.—Small frogs, males 15.8-20.1 mm. snout-vent length, single
female 18.5 mm.; vocal slits present in males; finger tips widely
expanded; first finger shorter than second; inner metatarsal tubercle
about three times size of outer; skin of dorsum shagreened, that of
venter smooth; tympanum concealed; pale cream in preservative with dark
brown spots; limbs banded; bands on forearm and thigh poorly developed
or absent; interorbital bar absent.
Remarks.—The tympanum is concealed in S. modestus, S.
nivocolimae, S. pallidus, S. teretistes, and to a lesser degree in
S. interorbitalis. However, if the specimen is permitted to dry
slightly, the annulus tympanicus becomes visible through the skin and a
tympanum/eye ratio can be computed.
One of the few cases of sympatry within the genus Syrrhophus involves
this species; modestus and nivocolimae are known to be sympatric at
one locality in southwestern Jalisco, México.
Duellman (1958) used the trinomial for this population and named a new
subspecies, pallidus, from Nayarit. I consider pallidus to be
specifically distinct from modestus because there is no evidence of
genetic exchange, and there is no overlap in the distinguishing
morphological features. I do consider the two populations to be closely
related but feel the inter-relationships between modestus, pallidus,
nivocolimae, and teretistes are more complex than would be indicated
by the use of trinomials. The sympatric occurrence of modestus and
nivocolimae is significant; morphologically, they might otherwise be
regarded as subspecies. Although allopatric, similar arguments could be
advanced for
[Pg 38]
the morphologically similar pallidus and teretistes.
The four are here afforded species rank since morphological similarity
and allopatry are not sufficient grounds for the assumption of genetic
exchange.
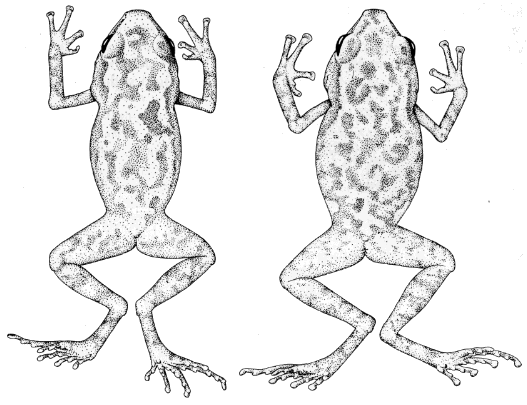
pallidus (right, UMMZ 115453). ×2.2.
Etymology.—Latin, meaning unassuming, modest, in reference to the
small size of the species.
Distribution.—Low elevations (up to 700 meters) in the lowlands and
foothills of Colima and southwestern Jalisco, México (Fig. 20).
Specimens examined.—(14). MÉXICO, Colima: Hda. Paso del Río, FMNH
100048 (holotype), 100167, 100299, UIMNH 13300, UMMZ 110877 (2), USNM
139729; 7.2 km. SW Tecolapa, UMMZ 115477 (4); Jalisco: 17.6 km. SW
Autlan, 606 m., KU 102627; 3.2 km. N La Resolana, UMMZ 102100; Bahía
Tenacatita, UMMZ 84264.
[Holotype.—LACM 3200, from Nevado de Colima (6 airline miles west of
Atenquique), Jalisco, México, 7800 feet; collected on July 20, 1964, by
Robert G. Webb].
Diagnosis.—Small frogs, males 18.5-21.1 mm. snout-vent length, only
known female 24.1 mm. snout-vent; vocal slits present in males; finger
tips widely expanded; first finger shorter than second; inner metatarsal
tubercle about three times size of outer; skin of dorsum warty, that of
venter smooth; tympanum concealed, its diameter 30.0-39.3 per cent that
of eye in males; mid-dorsal brown band from interorbital bar to anus;
bands on limbs narrow, dark bands
[Pg 39]
less than one-half width of light
bands, upper arm not banded; narrow interorbital light bar.
Remarks.—This species is closely related to S. modestus and differs
in color pattern and degree of wartiness of the skin. Dixon and Webb
(1966) held that nivocolimae had no close relatives, but the condition
of the tympanum, size, nature of the outer palmar tubercle, relative
sizes of the metatarsal tubercles, and shape and size of the digital
pads all point to a close relationship between S. modestus, S.
nivocolimae, and S. pallidus.
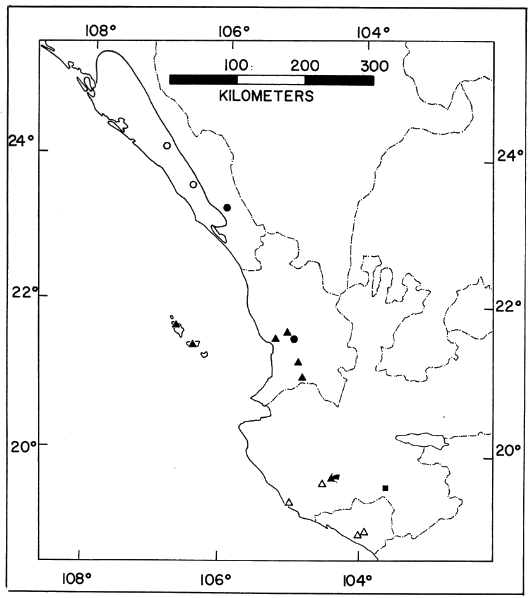
interorbitalis (open circles), teretistes (solid circles),
modestus (open triangles), pallidus (solid triangles) and
nivocolimae (square). Arrow indicates locality of sympatry between
modestus and nivocolimae. Solid line about the localities for
interorbitalis is a range estimate based on call records and specimens
examined.
Dixon and Webb (1966) reported that S. nivocolimae has a large
tympanum (50.0-59.0 per cent diameter of eye). However, my examination of
the type series and several other specimens from Jalisco reveals that the
[Pg 40]
largest tympanum/eye ratio is 39.3 per cent. Therefore, the
tympanum/eye ratio in S. nivocolimae is in agreement with those for
S. modestus, S. pallidus, and S. teretistes
(Table 6).
Etymology.—niv, Latin, and Colima (Nevado de), meaning high on the
volcano, in reference to the higher distribution of this species (around
2000 meters) than other members of the group.
Distribution.—Known from southwestern Jalisco, México, at moderate to
high elevations (600-2400 meters).
Specimens examined.—(48) MÉXICO, Jalisco: 17.6 km. SW Autlán, 606
m., KU 102626, 102631; 6.4 km. W Atenquique, 2060 m., KU 102628-30,
102632; 8 km. W Atenquique, 1970 m., LACM 3210-12; 9.6 km. W Atenquique,
2360 m., LACM 3200 (holotype), 3201-09; 14.5 km. W Atenquique, 2000 m.,
LACM 25424-36, 25439-41, 25446; 15 km. W Atenquique, LACM 37044-46,
37244-47; 16 km. W Atenquique, 2105 m., LACM 25443-45; 17 km. W
Atenquique, 2180 m., LACM 25442.
[Holotype.—UMMZ 115452, from San Blas, Nayarit, México, sea level;
collected on August 13, 1956, by William E. and Ann S. Duellman].
Zweifel, 1960:86-88, 91, 93-94, 118, 120-22. Gorham, 1966:166.
Diagnosis.—Small frogs, males 17.9-19.3 mm. snout-vent length; vocal
slits in males; finger tips widely expanded; first finger shorter than
second; inner metatarsal tubercle about three times size of outer; skin
of dorsum shagreened, that of venter smooth; tympanum concealed, its
diameter 27.0-35.6 per cent of eye in males; ground color cream
vermiculated with brown, upper arm and thigh lacking, or with few,
indistinct, bands; interorbital bar absent.
Remarks.—Considerable debate has been waged relative to the value of
subspecies and to the reasons for recognizing distinct disjunct
populations as species versus subspecies. Lacking evidence of genetic
exchange, I prefer to retain disjunct populations that are distinctive
as species.
All known specimens of pallidus can be separated from those of
modestus by color pattern. The two nominal species exhibit overlap in
proportions but the same can be said about nearly every species of
Syrrhophus; therefore, overlap in proportions can be disregarded in
assessing specific versus subspecific rank. Until contrary evidence is
forthcoming, I consider the disjunct populations heretofore held to be
subspecies of modestus to be specifically distinct. The specimens of
the disjunct population of pallidus on the Tres Marias do not differ
from the mainland population in Nayarit. This evidence, though perhaps
secondary, supports my contention that two species should be recognized.
Etymology.—Latin, in reference to the pale ground color in comparison
with that of S. modestus.
Distribution.—Low elevations in coastal Nayarit and on Islas Tres
Marias (Fig. 20).
Specimens examined.—(12) MÉXICO, Nayarit: 18.8 mi. NW Ahuacatlán,
UIMNH 7808; San Blas, UMMZ 115452 (holotype), 115453-57; 17 km. NE San
Blas, 150 m., MSU 5085; 12.8 km. E San Blas, UIMNH 71979; 31 km. E San
Blas, UIMNH 71978; 13.5 km. N Tepic, UIMNH 71980-81.
[Holotype.—UMMZ 115451, from 4.8 km. NW Tepic, Nayarit, México, 840 m.;
collected on August 12, 1956, by William E. Duellman]. Gorham, 1966:167.
Diagnosis.—Medium-sized frogs, males 19.2-23.2 mm. snout-vent length,
single known female 24.8 mm. snout-vent; vocal slits in males; finger
tips widely expanded; first finger shorter than second; inner metatarsal
tubercle about three times size of outer; skin of dorsum shagreened,
that of venter smooth; tympanum partially concealed, its diameter
28.6-43.8 per cent of eye in males; ground color brown vermiculated with
dark brown to nearly black; upper arm and thigh banded; interorbital
light bar absent.
Remarks.—S. teretistes appears to be most closely related to S.
pallidus; I consider it to be an upland derivative of pallidus.
Morphologically, the differences between the two are few, but lacking
evidence of genetic exchange they are retained as species.
Etymology.—Greek, in reference to the whistle-like nature of the
call.
Distribution.—Moderate elevations (840-1200 meters) in the Sierra
Occidental of Nayarit, Sinaloa, and Durango, México (Fig. 20).
Specimens examined.—(13) MÉXICO, Nayarit: 4.8 km. NW Tepic, 840 m.,
UMMZ 115451 (holotype). Sinaloa: Santa Lucía, 1090 m., KU 75263-72; 1
km. NE Santa Lucía, 1156 m., KU 78257; 2.2 km. NE Santa Lucía, 1156 m.,
KU 78258.
There are relatively few clear-cut morphological differences among the
fourteen species now assigned to Syrrhophus. The majority of the
species are allopatric and differ primarily in color patterns. Sympatric
occurrence serves as an indicator of specific distinctness and is one of
the more practical tests of species validity when cross-breeding
experiments are not possible. Two cases of sympatric occurrence are
known for the species of Syrrhophus in western México: modestus and
nivocolimae are sympatric in southern Jalisco and pipilans nebulosus
and rubrimaculatus are sympatric in southeastern Chiapas. In eastern
México, longipes and verrucipes are sympatric in southern Hidalgo,
and longipes is sympatric with cystignathoides, dennisi, and
guttilatus in southern Tamaulipas. Syrrhophus cystignathoides and
leprus are apparently sympatric in central Veracruz.
Subspecific assignments have been made only when there is evidence of
intergradation. The sympatric occurrence of morphologically similar
species in this genus has led me to adopt a conservative approach to the
degree of difference philosophy. I have therefore recognized all
morphologically distinct allopatric populations as species.
Tomodactylus (hatching). Black areas are zones of intergeneric
sympatry.
Widths of the columns are proportional to the numbers of species at a
given altitude; narrowest width equals one species.
Syrrhophus is closely allied to another Mexican leptodactylid genus,
Tomodactylus, which was revised by Dixon (1957), who along with
numerous other authors noted the close relationship between the two
genera. There is an almost complete lack of sympatry between the two
genera; in very few places in México do they coexist (Fig. 21).
Tomodactylus has its greatest diversity in the Cordillera Volcánica
and Sierra Madre del Sur, whereas Syrrhophus reaches its greatest
diversity in the Sierra Madre Oriental and eastern foothills. The
species of both genera are about the same size and presumably have
similar requirements insofar as food, breeding sites, and habitat
selection.
Four cases of intergeneric sympatry are known for the two genera: 1)
the Chilpancingo region of Guerrero, 2) the lowlands of Colima and the
mountains just inland in Jalisco, 3) the lowlands of central Nayarit,
and 4) the Sierra Madre Occidental on the Durango-Sinaloan border. The
apparent sympatry in the Chilpancingo region involves four species: S.
pipilans, T. albolabris, T. dilatus, and T. nitidus. Of the four,
T. dilatus appears to be completely allopatric in that it occurs at
higher altitudes (above 2000 meters), whereas the other three occur
below 1800 meters in the region (Davis and Dixon, 1965). In the
Colima-Jalisco region, Tomodactylus tends
[Pg 43]
to occur higher (Dixon and Webb, 1966) than some of the Syrrhophus,
but one subspecies of Tomodactylus nitidus is a lowland frog,
occurring sympatrically with the lowland Syrrhophus modestus. A
similar situation is observed in Nayarit; the lowland Tomodactylus
occurs sympatrically with the small Syrrhophus pallidus. In both cases
the Syrrhophus is smaller than the Tomodactylus.
Frogs of the genus Syrrhophus tend to occur at lower elevations than
do their close relatives of the genus Tomodactylus (Fig. 22). This
generalization is complicated by the occurrence in the Sierra Madre
Oriental in relatively high altitude Syrrhophus (up to 2000 m.) and
the occurrence in Michoacán of low altitude Tomodactylus (to sea
level). There are no Tomodactylus in the Sierra Madre Oriental,
whereas the genus Syrrhophus is represented in the lowlands of western
México (modestus group). Syrrhophus and Tomodactylus exhibit
essentially parapatric distributions. The two genera as now composed can
be characterized as low to moderate elevation frogs (Syrrhophus) and
moderate to intermediate elevation frogs (Tomodactylus).
Survey, pp. 1-35, pls. 1-41.
Club, 8:81-83.
Zool., 96:59-214.
1888, pt. 2:204-06.
So. California Acad. Sci., 61:193-203.
Boston. 366 pp.
Anura. J. Acad. Nat. Sci. Philadelphia, n. ser., 6:67-112.
Amer. Philos. Soc., 17:85-98.
Amer. Philos. Soc., 18:261-77.
Herpetologica, 13:145-47.
Imprenta de Ricardo Rodriquez Romo. Aguascalientes. 40 pp.
in Mexico. Texas J. Sci., 9:379-409.
Cont. Sc., Los Angeles Co. Mus., 102:1-5.
Occ. Pap. Mus. Zool. Univ. Michigan, 594:1-15.
Mexico. Univ. Kansas Publs. Mus. Nat. Hist., 13:19-72.
genus Syrrhophus, with a description of a new species. Copeia,
(1):48-58.
12:201-15.
Soc. London, 1905, pt. 2:191-244.
Lief, 85:1-222.
76 pls. Syrrhophus section dated 1900.
Bull. U.S. Natl. Mus., 160.
Herpetologica, 12:161-65.
(Amphibia: Leptodactylidae). Copeia, (3):580-81.
Texas and Florida Herpetologica. 20:141-42.
under the plenary powers. I. N. (S). Bull. Zool. Nomencl.,
24:313-15.
Nat. Hist., 17:503-15.
region, Tamaulipas, Mexico. Misc. Publs. Mus. Zool. Univ. Michigan,
101:1-102.
Terrell County, Texas. Texas J. Sci., 2:543-62.
Bull. Florida State Mus., 9:77-130.
Monatsb. k. k. Preuss. Akad. Wiss. Berlin, 1870:641-52.
Mus. Nat. Hist., zool. ser., 29:75-96.
Sci., 37:408-12.
U.S. Natl. Mus., 194:1-118.
Soc. Washington, 28:131-32.
Zool. Club, 18:13-16.
89:43-47, 1 pl.
53:95-98, 1 pl.
the Walter Rathbone Bacon Traveling Scholarship. Proc. U.S.
Natl. Mus., 95:521-613.
Oklahoma and Texas. Southwest. Nat., 5:66-70.
of specimens in U.S. National Museum. Bull. U.S. Natl. Mus.,
24:1-249.
to Western Mexico. 9. Herpetology of the Tres Marias
Islands. Bull. Amer. Mus. Nat. Hist., 119:81-128.
Although Syrrhophus marnocki and Syrrhophus marnockii
both appear in this text, a literature search shows that both
spellings have been used and the two instances where there is only
one “i” at the end are in reference to priviously published names.
Therefore, they were left as is. With the exception of the list below
and a number of silent corrections, the text presented is that of the
original printed version. The original cover was modified to
include graphics from the article.
| Page | Correction | |
| 3 | otherwse => otherwise | |
| 5 | poltypic => polytypic | |
| 12 | interorbtal => interorbital | |
| 14 | neublosus => nebulosus | |
| 16 | Cuidad => Ciudad | |
| 16 | 1946-170 => 1946:170 | |
| 22 | rubrimacultaus => rubrimaculatus | |
| 27 | resemblence => resemblance |